

Fish Bite Force
Gape-specific bite force and prey-size specific predator performance in the snail-eating black carp
- Brown University, Providence, RI, USA
- The University of California at Davis, Davis, CA, USA
*Author for correspondence: Nicholas J. Gidmark | Published article | Press release
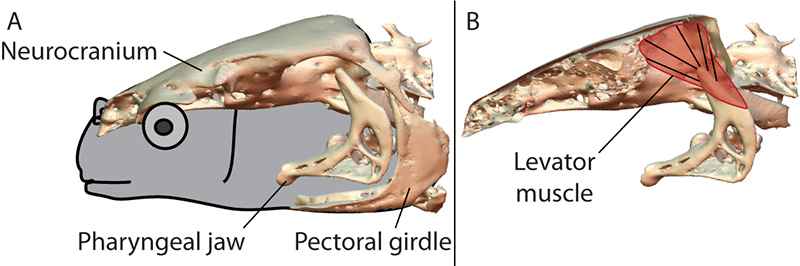
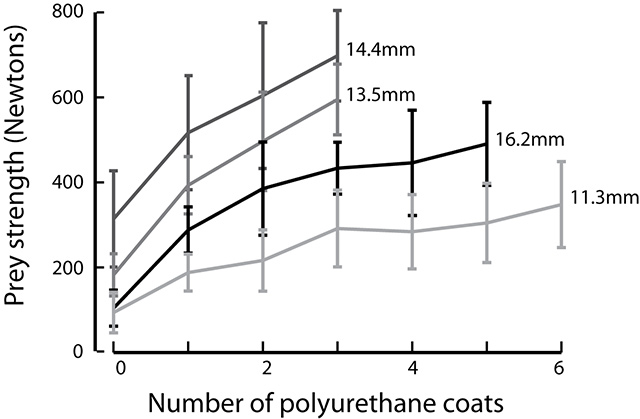
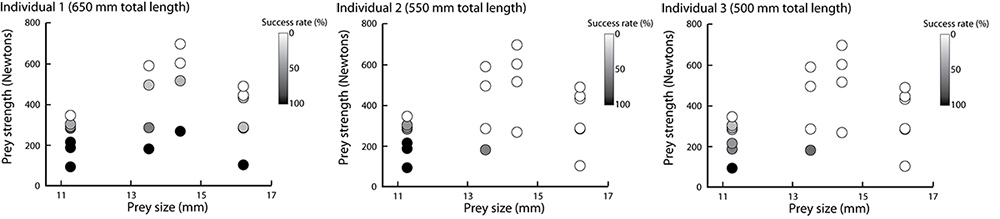
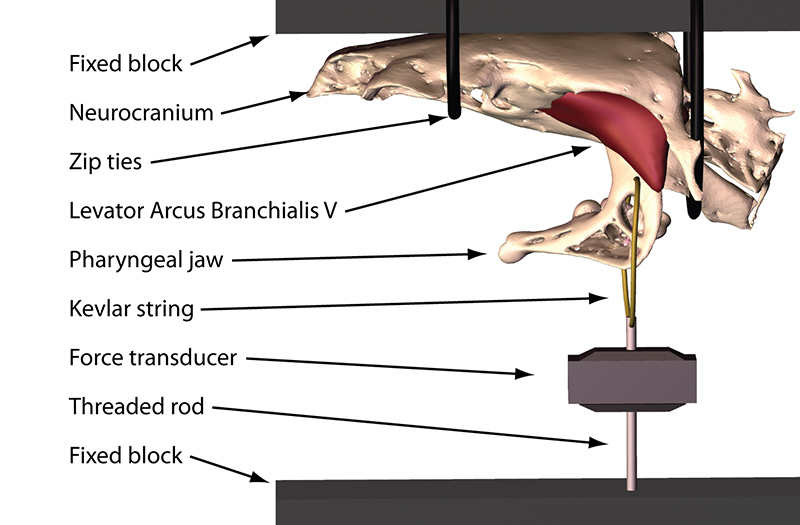
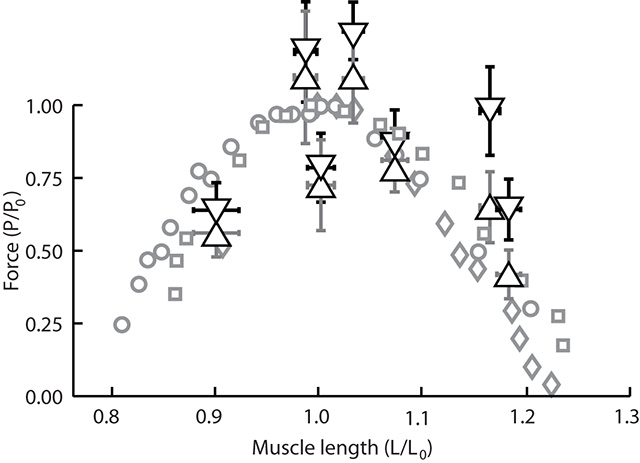
In this study we used X-ray Reconstruction of Moving Morphology (XROMM) to examine biting performance in a snail-eating carp species, the black carp (Mylopharyngodon piceus). Instead of using its oral jaws for crushing, black carp use the pharyngeal jaw apparatus, in the back of the throat (
Press Coverage
Size of lunch dictates force of crunch
Posted Feb. 12, 2013 on news.brown.edu. Story by David Orenstein.
Additional coverage by: news.science360.gov | eurekalert.org | sciencedaily.com | ria.ru/science | scienceblog.com | redorbit.com | nsf.gov/news | sciencenewsline.com | esciencenews.com | phys.org/news
Reference
Gidmark, N.J., N. Konow, E. LoPresti, and E.L. Brainerd. (2013). Bite force is limited by the force-length relationship of skeletal muscle in black carp, Mylopharyngodon piceus. Biology Letters, 9:20121181. Published article. Press release.
